 |
 Как обустроить мансарду?  Как создать искусственный водоем?  Как наладить теплоизоляцию?  Как сделать стяжку пола?  Как выбрать теплый пол?  Зачем нужны фасадные системы?  Что может получиться из балкона? |
Главная страница » Энциклопедия строителя
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5]
страница - 0
Квази-трехмерная модель свободно растущего дерева
В.В. Галицкий (galvv@ibbp.psn.ru)
Институт физико-химических и биологических проблем почвоведения РАН
Введение
Понятие - свободно растущее растение - было использовано в работах (Галицкий, 1999; 2000; Galitskii, 2003) в качестве одного из основных элементов двухмерных моделей растения и сообщества растений, предназначенных, в первую очередь, для изучения влияния свойств индивидуальных растений, составляющих сообщество, правил их взаимодействия и локальной структуры сообщества на его динамику или иными словами - проблемы "микроскопической" обусловленности макроскопического поведения сообщества. Свободно растущее растение по определению не испытывает влияния конкуренции за ресурсы со стороны других растений сообщества, но находится в экологических условиях, создаваемых сообществом. В упомянутых моделях оно является некоторым идеалом, относительно которого рассматривалось поведение (угнетаемого) растения. Оно позволяет выделить и описать процесс собственно конкуренции из множества разнообразных процессов, в которых участвует и влияние которых испытывает растение в сообществе.
Сходные понятия - "отдельно", "изолированно", "открыто растущее" растение - иногда рассматриваются в литературе (Сукачев, 1972; Dale et al., 1985). Различие состоит в том, что там обсуждаются конкретные, существующие в природе объекты - растения, достаточно удаленные пространственно от других растений и потому не испытывающие конкуренции, но и находящиеся в условиях (в частности, микроклиматических), отличных, вообще говоря, от условий, имеющихся в сообществе взаимодействующих растений. Используемый же нами (умозрительный) объект - свободно растущее растение - находится в условиях сообщества, не испытывая конкуренции. По сути, характеристики свободно растущего растения могут быть использованы (теоретически, во всяком случае) в качестве такой меры "нормальной производительности данных условий местопроизрастания" или бонитета территории (Сукачев, 1972, с. 93), которая (мера) не была бы отягощена артефактами, связанными с влиянием структуры и истории конкретного сообщества.
В двухмерном случае в качестве характеристик свободно растущего растения (его имитационной модели) были использованы две функции от возраста растения: AF(T) -площадь, необходимая растению для свободного роста в возрасте T и BF(T) -соответствующая биомасса. Под биомассой понимается физиологически активная часть полной массы растения (Галицкий, 1999; Galitskii, 2003). Форма площади в случае изотропной территории - круг увеличивающегося с возрастом радиуса. Получение этих функций, связывающих физиологию, морфологию и т.п. внутренние аспекты жизнедеятельности растительного организма с экологическими условиями его существования, исключая конкуренционное взаимодействие с соседними растениями, должно представлять собой предмет других моделей и лежит вне рассматриваемой здесь задачи. Наша цель - использовать эти функции и для рассматриваемой далее квазитрехмерной модели.
И. А. Полетаев (1966) с помощью несложной модели, основанной на вполне разумном предположении, показал, что рост дерева в высоту физически ограничен необходимостью подъема влаги и может быть описан как
H(T) = H0 th(TA).(1)
В соответствии с этим результатом в двухмерных моделях (Галицкий, 1999; Galitskii, 2003) в качестве биомассы свободно растущего дерева было использовано
BP (T) = B0 th1 (T/A1),(2)
где H0, A1; B0 - размерные параметры и ц - аллометрический параметр, в случае изометрии равный 2 при B-типе роста (Галицкий, 1999).
Двухмерное представление (моделирование) трехмерных растений и их сообществ может рассматриваться в соответствии с "принципом минимального угла зрения" (Галицкий, Тюрюканов, 2001) как некоторый этап на пути построения ряда постепенно усложняющихся моделей объекта и, соответственно, имеет свои пределы качественной и количественной адекватности моделируемому объекту. Примеры этого можно найти в двухмерном модельном анализе правила -3/2 (Галицкий, 1998) и эффекта не монотонности конкуренции в однородных древесных культурах (Галицкий, Абатуров, 1996; Галицкий, 2003). В качестве следующего узла в некотором ряду моделей может быть рассмотрена трехмерная модель растения.
Ниже рассматривается некоторая детализация двухмерной модели свободно растущего дерева - квази-трехмерная модель, предназначенная для использования в качестве элемента общих квази-трехмерных моделей дерева, из которых строятся модели сообщества деревьев. Трехмерность для наших целей достаточно просто и наглядно (по крайней мере, для хвойных деревьев бореальных местообитаний) вводится путем выделения в биомассе дерева составляющих ее секций, каждая из которых может быть сопоставлена с суммарной биомассой одновозрастных ветвей, относящихся к одному узлу (с ветвями) ствола дерева, и соответствующего междоузлия. Такой морфологический способ выделения секций биомассы не единственный, но как будет видно, вполне конструктивный. Использование префикса квази- в характеристике модели связано с тем, что в описываемой модели в действительности нет полноценного описания третьего измерения, т.к. рост дерева в высоту (как и периодичность появления секций) не определен внутри модели и не связан с ее функционированием, а задается снаружи, что впрочем на начальном этапе построения и анализа модели не существенно.
Динамика биомассы секции
В рамках общей концепции движения от рассмотрения объекта под "минимальным углом зрения"к постепенной детализации представлений об его устройстве, вводя в рассмотрение новую деталь - секцию биомассы дерева, попытаемся установить ее соотношение с соответствующим объектом предыдущего уровня - биомассой всего дерева.
Начиная с момента появления каждого новой /-ой секции (стволовые мутовка и междоузлие) дерево начинает реализовывать новую (которая может быть той же самой, что была у предыдущей секции) в определенной степени виртуальную динамику биомассы BPji(T-T) верхней части дерева, начинающейся с этой секции (BPji(x)=0, x<0). С нулевой секции начинает реализовываться BPj0(T) - виртуальная динамика биомассы всего дерева. Каждая из этих динамик действительна вплоть до появления новой следующей секции, а затем ее можно считать действительной в части, относящейся к собственно данной секции и виртуальной для
следующей секции, поскольку следующая секция реализует аналогичным образом свою динамику.
Очевидно, что биомасса всего дерева есть сумма биомасс всех его секций, имеющихся на данный момент времени. Предположим, что с момента появления на макушке дерева новой i-ой секции динамика части биомассы дерева, начинающейся с этой секции, описывается той же самой функцией свободного роста BP(T), что и для всех предыдущих секций дерева (в этом случае совпадающей с функцией для всего дерева), но сдвинутой на момент времени появления i-ой секции. Вплоть до появления следующей (1+1)-ой секции эта функция описывает также и динамику биомассы i-ой секции. То же самое можно сказать и о следующем (1+1)-ой секции, но уже с учетом сдвига на момент появления этой секции. После появления (i+1)-ой секции динамика биомассы i-ой секции может быть вычислена как разность биомассы дерева, начиная с этой секции, и биомассы дерева, начиная с (1+1)-ой секции.
Таким образом, если секции биомассы дерева появляются с шагом по времени AT, то секция, появившаяся в момент Ti будет иметь зависимость биомассы от времени
bP/T) = BP/T-T) - BpI+](T-T- AT),(3)
где все функции от отрицательного аргумента равны нулю. Очевидно, что, просуммировав такие "парциальные" функции по всем имеющимся на данный момент секциям, мы получаем заданную функцию BP(T) для всего дерева.
Из выражения (3) следует, что вид зависимости от времени биомассы собственно секции bP,i(T) может качественно отличаться от зависимости BP(T) для всего дерева. Если полагать, что все дерево имеет ^-образную функцию BP(T) (B-тип роста, например, B0 thP(T/A\), Галицкий, 1999), то секция будет иметь колоколообразную зависимость биомассы от
0.008
0.004
0.000
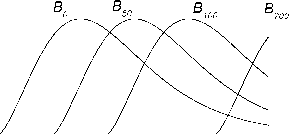
050100150 200 250
T
Рис. 1. Динамика биомассы некоторых секций
времени, т. е. со временем биомасса секции (биомасса ветвей данного узла и соответствующего междоузлия) должна отмирать. Таким образом, нижние более старые ветви дерева могут отмирать не только из-за конкуренции и других биологических обстоятельств, но и по "математическим" причинам.
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5]